The Significance of the Flocculus in Archosauria
With advancements in computed tomography (CT) scanning comes an increased understanding of the internal structures preserved in extant and extinct animals, providing a non-destructive way of peering into the bones and revealing their secrets. Along with this month’s publication on extensive rostral neurovasculature in the carcharodontosaurian Neovenator, comes the CT scanning of the braincase of a distantly related theropod, Viavenator exxoni. What interested me in this paper on Viavenator, discovered last year in 84 million year old Patagonian rocks, was the preservation and description of the flocculus, a small area of the brain that we will return to in more depth below, and its importance in animal behaviour.
A (Very) Brief and Unflattering Look Into Abelisaurid Palaeobiology
“Peculiar” is a word one could describe the abelisaurids, the clade of reptiles Viavenator shares its ancestry with. The arms in this clade are excessively short, bordering on vestigial, with the bones of the palm (metacarpals) attaching directly to the humerus. The fingers bear tiny claws, or are devoid of them completely in more derived forms, and the elbow joint was immobile so that the arms are held straight at all times, making the famously puny-armed tyrannosaurs positively dexterous in comparison (interestingly, mutation of the HOXA11 and HOXD11 genes has been suggested as the reason for such reduced forelimb anatomy in abelisaurids, although a total loss of these homeobox genes is unlikely due to the associated urogenital deformities). The majority of this clade are from the southern hemisphere, formerly known as Gondwana, although rarer European forms exist.
The skulls in the slow-growing abelisaurids are short, tall and wide, providing a box-like appearance that lacks the tapered snout seen in other theropods, with the bone texture often described as “rugose” or “sculpted”. The cervical vertebrae are robust, suggesting a heavily-muscled neck, possibly hinting at a different predation style to the puncture-pull method of tyrannosaurs or the slash-tear arsenal of allosauroids, perhaps preferring to engage in prolonged skull-prey contact. To add to the weirdness, the axial skeleton (the bones that make up the neck, spine and, of particular interest, the tail) is stiffened in this group, which may have allowed further development of musculature associated with running, perhaps allowing them to outcompete contemporaneous theropods.
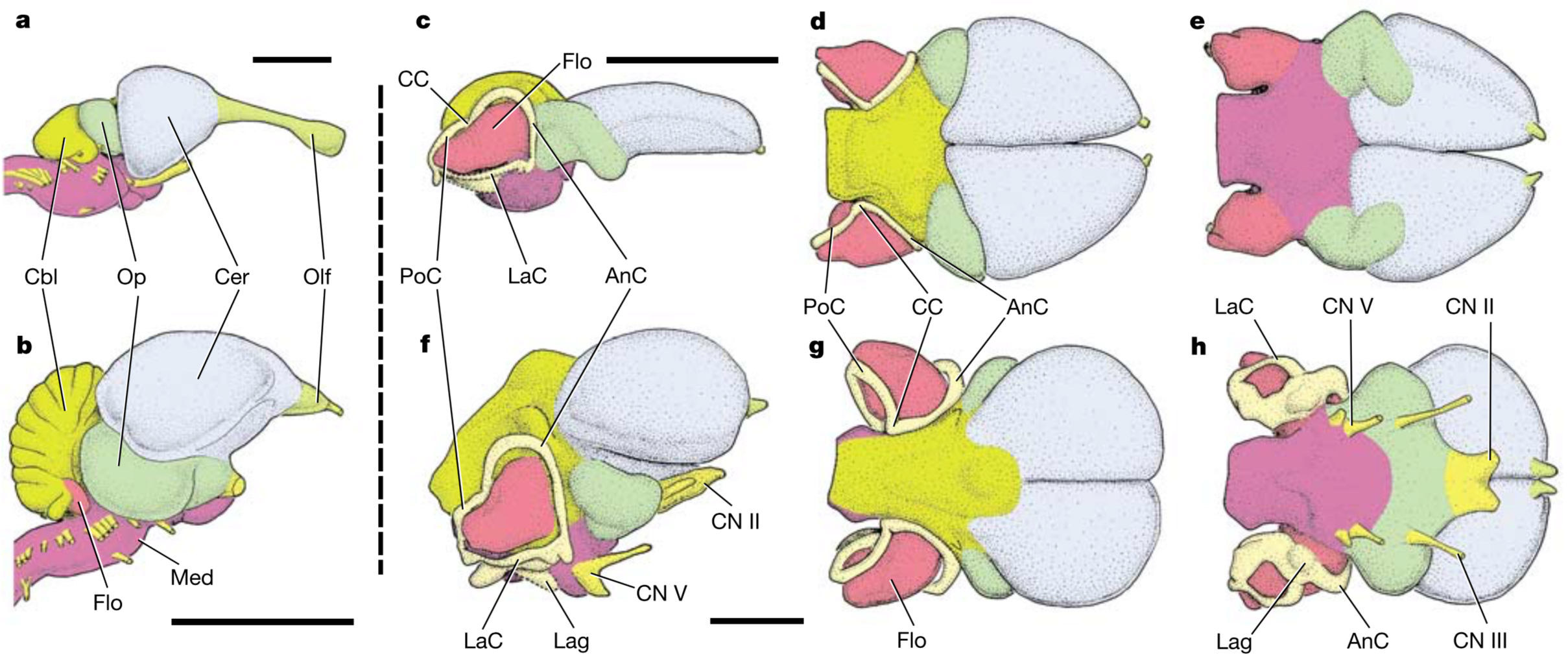
The braincase (left) and virtual endocast (right) of Viavenator, showing the flocculus (floc) within the semicircular canals of the inner ear (coloured in pink and denoted as ie). Credit: Paulina-Carbajal and Filippi, 2017.
Inferring Viavenator’s Sensory Capabilities
The CT results indicated that Viavenator and other abelisaurids shared a characteristic neuroanatomy, with Paulina-Carbajal and Filippi writing similarities in “general shape and proportion of the regions of the brain, large olfactory tract and bulbs, similar inner ear morphology…” were found throughout this clade. These generalities were expanded to ceratosaurs and other basal tetanurans (theropods more derived than the aforementioned ceratosaurs) as a whole, which encompassed characters including “large dorsal longitudinal sinuses, small flocculi, poorly defined optic lobes, and cerebral hemispheres that do not project far laterally”. In terms of hearing capabilities, Viavenator demonstrates classic large dinosaur hearing ranges with a high-frequency limit below 3 kHz (humans can hear up to 20kHz).
Of particular interest to this blog post are the flocculi, which in birds and dinosaurs is housed in a small bony recess, making it easy to identify in CT-generated endocasts. Part of the oculomotor reflex within the cerebellum, the flocculus has been historically associated with flying and bipedalism, due to the close proximity to the semicircular canals that provide balance and the inherent need to be aware of one’s body in these unstable forms of locomotion. On the other side of the archosaur tree, it is not present in modern crocodilians, however it is found in several of their extinct cousins within the clade Pseudosuchia. This raises an interesting point as some members of this clade, such as aetosaurs, and loricatans, are in fact quadrupedal, implying that the presence of a flocculus is not strictly related to flight or walking on two legs.
The flocculi perform two important reflexes: the vestibulo-collic reflex (VCR), which acts to stabilise the head via impulses to the neck musculature, and the vestibulo-ocular reflex (VOR), which acts to maintain a stable image on the retina when the head is moving. This is particularly important in some modern archosaurs, such as in birds that engage in sophisticated aerial/aqautic manoeuvres (i.e., aerial/aquatic predators) or those prone to unstable low speed flying/hovering (e.g., hummingbirds), preventing blurring and allowing these animals to maintain a steady gaze on an object.
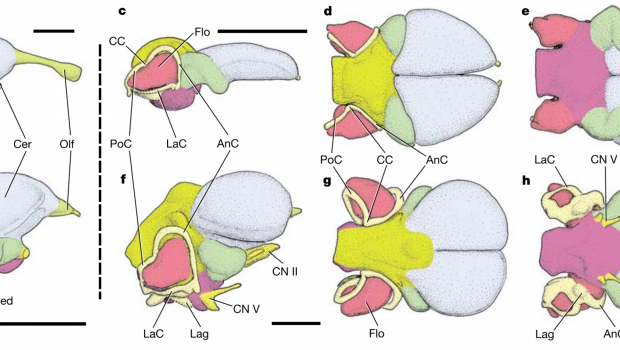
Returning to Viavenator (Fig. 1), the flocculi, described as small relative to other theropods or flying reptiles, are greatly expanded when viewed next to its African cousin Majungasaurus (Fig. 2). The authors believe this increase in volume allows Viavenator enhanced gaze stabilisation versus Majungasaurus, suggesting these South American forms rely on rapid head movements more so that their African relatives.
The endocast of three abelisaurids: the South American Viavenator (A, B) and Aucasaurus (D, E), and the African Majungasaurus (G, H), with the semicircular canals of the inner ear in dark grey and flocculus (floc) within them. Note the expansion of the flocculus in the two South American forms compared to the African representative. Credit: Paulina-Carbajal and Filippi, 2017.
The Floccular Fossil Record
As noted above, other non-avian theropods, including tyrannosaurs, have larger flocculi compared to these abelisaurids, which, when coupled with its presence in quadrupedal pseudosuchians, implies gaze stabilisation, rather than balance, was key to this expansion. It is easy to imagine the use of such an organ in predators like carnivorous theropods, as well as the use in fossil bird taxa, but what use does it have in the herbivorous individuals?
The therizinosaurs Erlikosaurus and Nothronychus, members of the coelurosauria group that includes tyrannosaurs and “raptors”, have relatively large floccular lobes, yet have become secondarily herbivorous. Proposed functions of gaze stabilisation in therizinosaurs range from territoriality to courtship, or more simply a remnant of their coelurosaurian heritage. However, if we couple this adaptation with the evidence of an enlarged forebrain in therizinosaurs, and the well-developed sense of smell, hearing and balance, it is possible this neurosensory complex may have been crucial in foraging, predator evasion and/or social complexity.
Several other prehistoric endocasts reveal floccular lobes, however its presence is not uniform among taxa. Joining the theropods are the armoured ankylosaurids, however the flocculus is not found in their close cousins the nodosaurids, nor is it declared present in endocasts from other ornithischians (bipedal or otherwise) bar several stegosaurs (Stegosaurus ungulatus, S. stenops, and Kentrosaurus aethiopicus). Interestingly, the presence of a floccular lobe in thyreophorans like ankylosaurids and stegosaurids, with their tail clubs and spikes unseen in the nodosaurids, may reflect the need for gaze stabilisation when actively using their tail weaponry.
Perhaps even more bizarrely is the presence of floccular lobes in a handful of species of sauropod, including the rebbachisaurid Nigersaurus, the dicraeosaurid Dicraeoesaurus and the titanosauriformes Giraffatitan. The reasons as to why they would have needed advanced gaze stabilisation mechanisms is not mentioned in the literature, nor how large it is compared to the taxa discussed above, although this could be an interesting topic to explore in the future.
The presence of flocculi in pterosaurs shouldn’t come to any surprise. Like the distantly-related birds, aerial maneuverability would have been key to many species, however unlike their avian cousins, pterosaurs did not evolve larger brains. Indeed, the buzzword for pterosaur evolution appears to be ‘weight reduction’, and expanding the brain was deemed an unnecessary burden. As a result, to deal with the complexities of flight, pterosaurs greatly expanded the inner ear and flocculus region, to the point that some species such as Rhamphorhynchus and Anhanguera, the flocculus has become larger than the optic lobe, and was a site of neural processing “unlike that seen in any extant animal”. Intriguingly, despite these differences, pterosaurs are no more adapted to an aerial lifestyle than birds, and one would expect convergence between the two due to shared ecological niches.
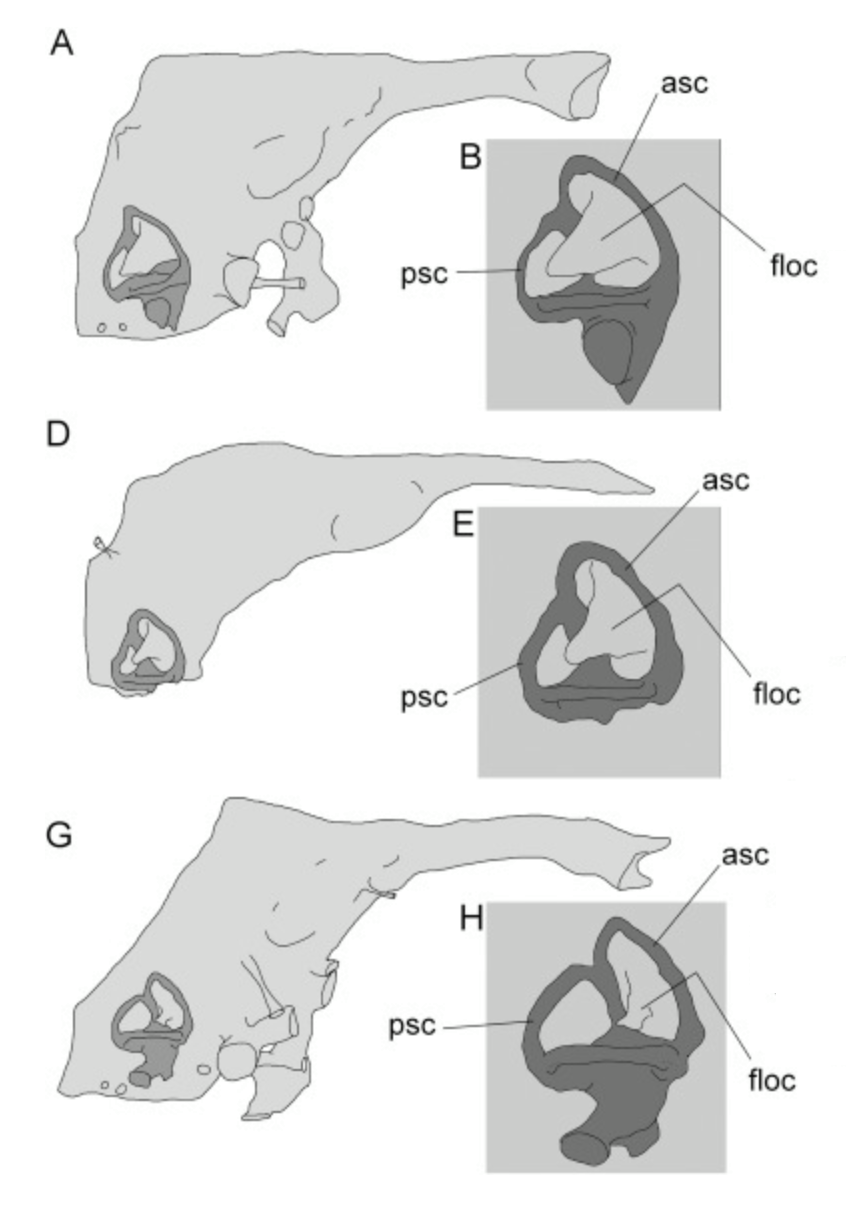
Endocasts of a, Brain of the crocodilian archosaur Alligator mississippiensis in right lateral view. b, Brain of the avian archosaur Columba livia (pigeon) in right lateral view. c–e, Endocast and osseous labyrinth of Rhamphorhynchus muensteri in right lateral (c), dorsal (d), and ventral (e) views. f–h, Same of Anhanguera santanae in right lateral (f), dorsal (g), and ventral (h) views. Note the expanded flocculus (Flo) in the pterosaurs, denoted in red, versus the green optic lobes (Op). Credit: Witmer et al., 2003.
Witmer and his colleagues suggested that this hugely expanded floccular region may have received proprioceptive and somatosensory information associated with the wing membrane (a highly-complex structure made of muscle and tendon, not just a layer of skin, that was unique to the clade). As a result, adjustments to the wing membrane would allow for changes in body position, thus rendering the gaze steady and uninterrupted.
Cranial Caveats
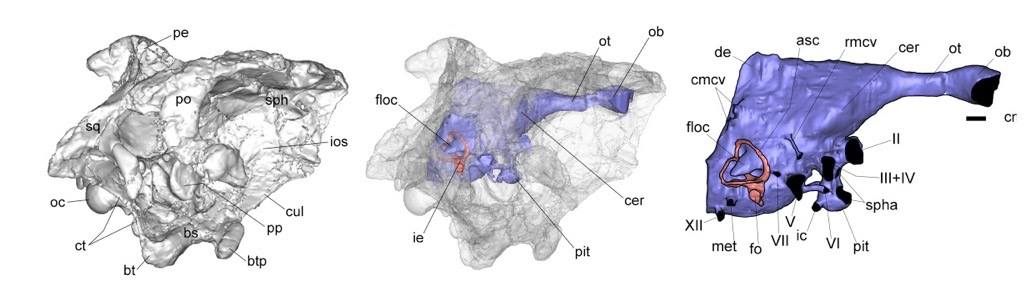
However, as is the way with the palaeontology, reconstructing the behaviour of extinct animals using osteological correlates may not accurately reflect their true behaviour. Indeed, a study of 60 extant bird species found no correlation between the size of the bony cavity that houses the flocculus, and flying ability.
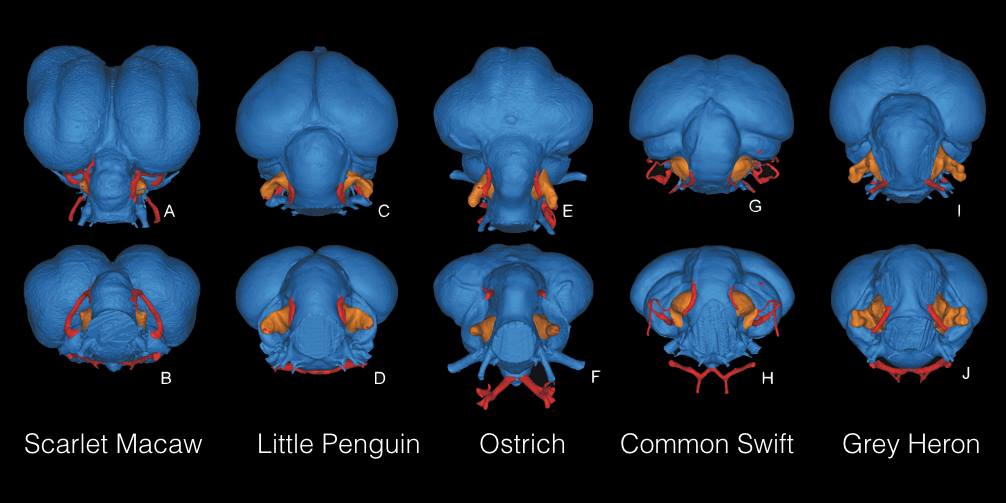
The five floccular types, shown in orange surrounded by the red semicircular canals, recovered in the study, the top row in dorsal view and the bottom row in caudal view. Credit: Walsh et al., 2013.
There are a multitude of possible reasons for this result, with arguments ranging from the improbable (i.e. that a significant portion of the nerves in the flocculus are hidden within the cerebellum), to the more problematic, and possibly more real, issue whereby a large proportion of the bony cavity houses blood vessels and not entirely neural tissue. This brings serious consequences for our ability to decipher extinct animal behaviour as we cannot estimate the neural volume in fossil endocasts. This is further exacerbated by the fact that many of the animals mentioned above are distantly-related to modern birds, and as such may have had vascular and/or neural tissue completely unknown to modern clades. As Walsh and colleagues write, “an extensive survey of vascular versus neural occupancy of the floccular fossa in living bird species is needed to test the nature and extent of this variability”.
Interestingly, a recent article in Scientific Reports explored the relationship between floccular fossa size and behavior of living animals, in order to assess whether such correlations could be attributed to extinct animals. Forty-seven mammalian species and fifty-nine bird species were micro-CT scanned to test 6 possible hypotheses regarding the fossae volume and its significance for ecological/behavioural traits. These hypotheses ranged from body mass, agility and feeding styles, to locomotion type and activity patterns.
Importantly, they note that many of the palaeontological studies on this subject are based on a double assumption: firstly, that brain endocasts reflect brain volume in the living organism (i.e. the brain occupies the whole brain cavity), and secondly, that large floccular lobe volumes in animals from various habitats are a result of adaptive pressure to make them more agile due to shared selective pressures to develop highly coordinated movements.
A lack of correlation was found in all but two of the six hypotheses tested, with the two significant results requiring further empirical testing to assess their validity. Both significant results occurred in the avian test subjects, with feeding/activity pattern vs floccular fossa size showing correlation despite high variability. Larger fossae size was seen in predators compared to occasional predators and gatherer species, which make sense seeing as visual accuracy is an important trait in these animals. It also supports the hypothesis that large fossae size (as a result of overall cerebellar growth) in carnivorous theropods is related to their evolution of pursuit predatory lifestyles. Furthermore, this already large fossae size may have been important in the evolution of flight in their avian descendants.
The authors concluded that their data “does not support floccular fossae size as a reliable proxy for inferring ecology and behaviour”, and the reason for this may lie within the cerebellum. This part of the brain is highly plastic and adaptable, with parts of the flocculus originally used for eye movement control could evolve to serve a new function, or that the fossae itself is occupied by parts of the enigmatic paraflocculus, which is used for different functions (i.e. in mammals, this tissue is capable of eye motor control in some species, but is also seen to influence arm movement control). This cerebellar plasticity, the authors note, is why assumptions regarding animal behaviour based on floccular fossa volumes should be approached with caution, at least until further empirical testing is conducted on the subject.
 Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Related Posts

Why did Sauropods grow so big? →

200 Years of Dinosaurs →

Modern Vs Archaic Mammals, a match-up decided by brains →













