
Episode 18: Trilobites
Trilobites are one of the most instantly recognisable groups of fossils. They were present from the very start of the Paleozoic and went on the fill a great number of ecological roles before going extinct at the Permo-Triassic mass extinction event 252 million years ago. They ranged from the very small to the very large, from the most basic appearance to the most elaborately ornamented.
We were lucky enough to get the opportunity to speak to Prof. Richard Fortey of the Natural History Museum, London, all about trilobite morphology and ecology.
Podcast: Download (Duration: 57:55 — 79.5MB)
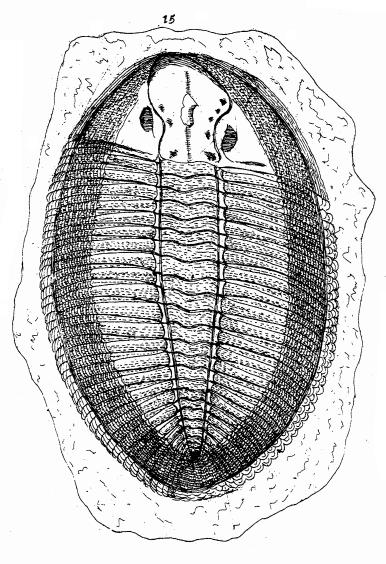
Trilobites were first figured in the 1698 volume of “The Philosophical Transactions of the Royal Society” (coincidentally the first scientific journal written in the English language), where the Rev. Edward Lhwyd discusses their affinity:
“The 15th [figured specimen] whereof we found great Plenty, must doubtless be referred to the Sceleton of some Flat Fish…”
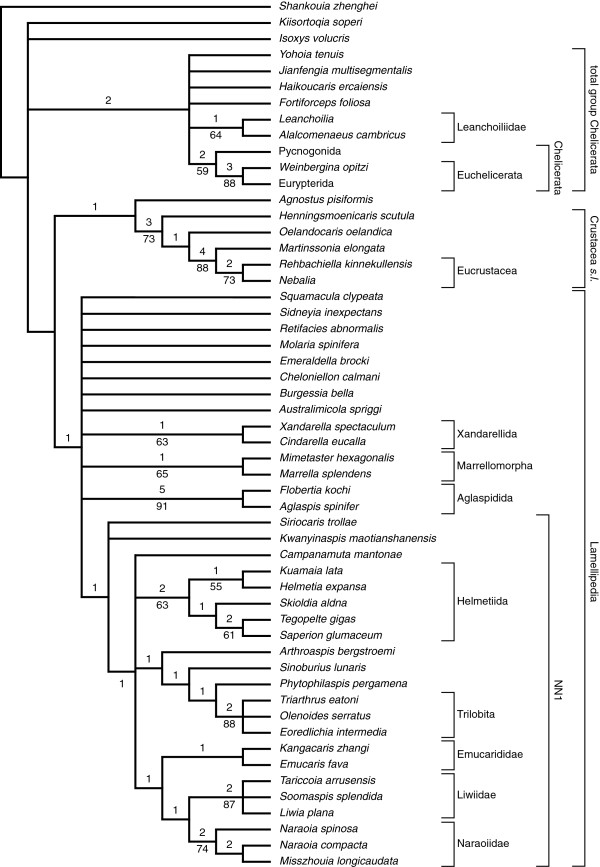
The true affinities of trilobites are still contested; the latest phylogenetic studies suggest trilobites, in this case as part of a group ‘Lamellipedia’, are sister-group to crustaceans. From Stein et. al. 2013.
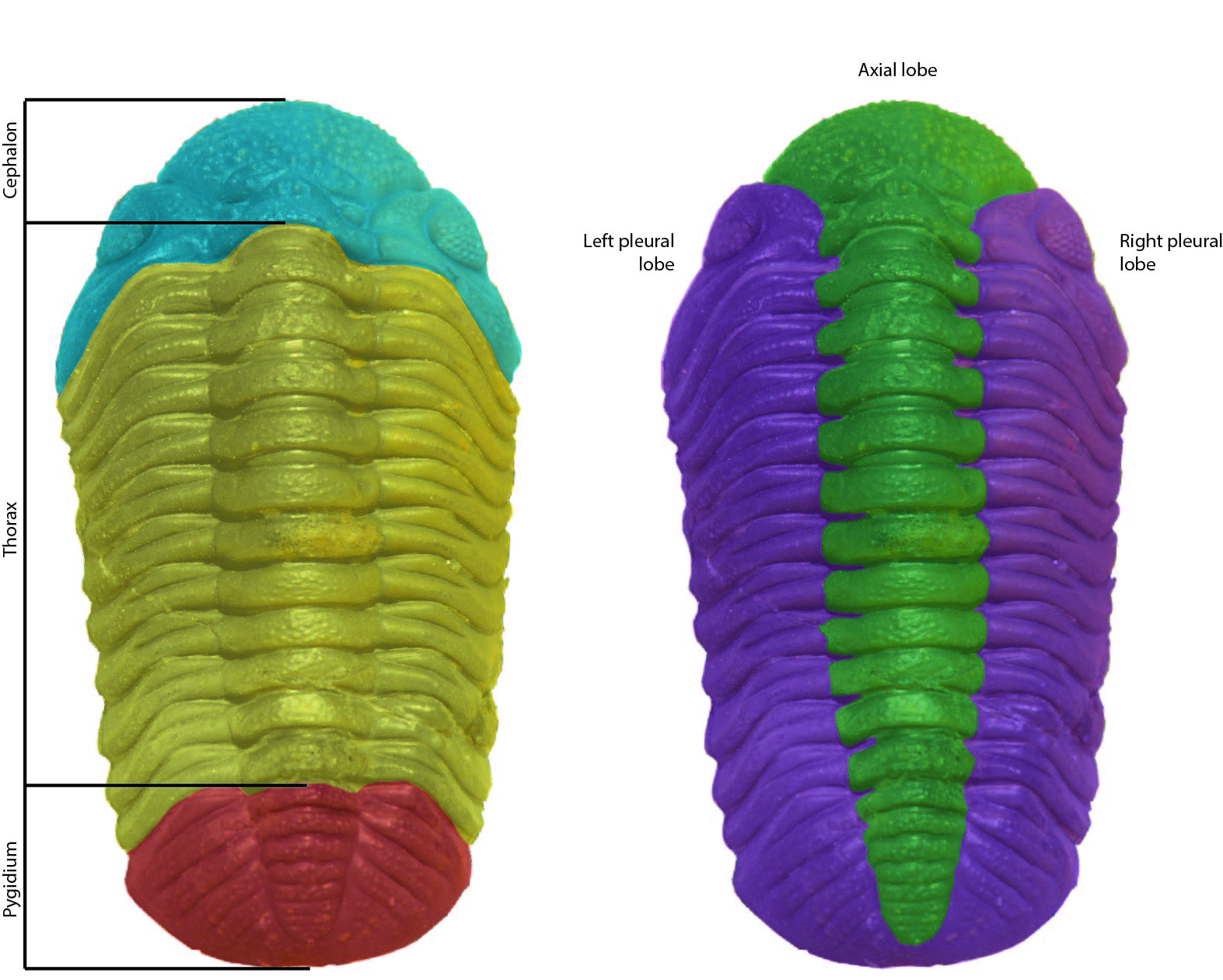
The name ‘trilobite’ means ‘three lobes’ (tri-lobe) and is often mistaken for their tagma; the cephalon (head), thorax (body/preabdomen) and pygidium (tail piece/ postabdomen). The name actually refers to the division of the animal into the left and right pleural lobes and the centrally positioned axial lobe.

Despite having such a simple appearance (dorsally at least), there is a great amount of variation that can be built into the number of segments, relative proportions of each tagma and location of spines. These small differences contribute to the recognition of approximately 17,000 individual species.

The physical characteristics of trilobites are often used to infer how each species lived; morphology to infer ecology. Dense pustulation (lumps and bumps) have been suggested to protect the carapace from abrasion. Feeding habits can also be inferred from the ventral feeding plate, the hypostome, and the glabella and their positioning relative to each other.

Other characteristics, such as these spines, served a less cryptic function.

Which becomes particularly apparent when the trilobite enrols.

Dicranurus monstrosus possesses large, ornate spines, which may potentially serve a purpose other than just protection.

Walliserops trifurcatus is a good example of how we can infer the ecology of an extinct animal from an unknown structure, even with no living relatives for comparison. For further discussion see Knell and Fortey 2005 or listen to Palaeocast episode 11.

Visual systems are also an important factor in determining ecology and also display evolution in form. This benthic (bottom dwelling) trilobite possesses holochroal eyes, positioned to look out across the sea floor. Copyright free image.

This trilobite possesses the more advanced schizochroal eye, this one expanded dorsally, to give an even better view of the sea floor. Modifications in other trilobites even include sun-shades to isolate the lenses from immediate light from above.

Some pelagic trilobites (those freely swimming in the ocean), such as this Symphysops, featured greatly enlarged eyes.

These two eyes were so large, that they met in the middle, forming one continuous visual field.

Other morphological adaptations include expanded and pitted cephalic fringes, as seen in the harpetids and trinucleids. The function of these fringes is still being argued, but its possible functions include sieving food out of sediment or for detecting small movements in the water, as a sort of ‘compound ear’.
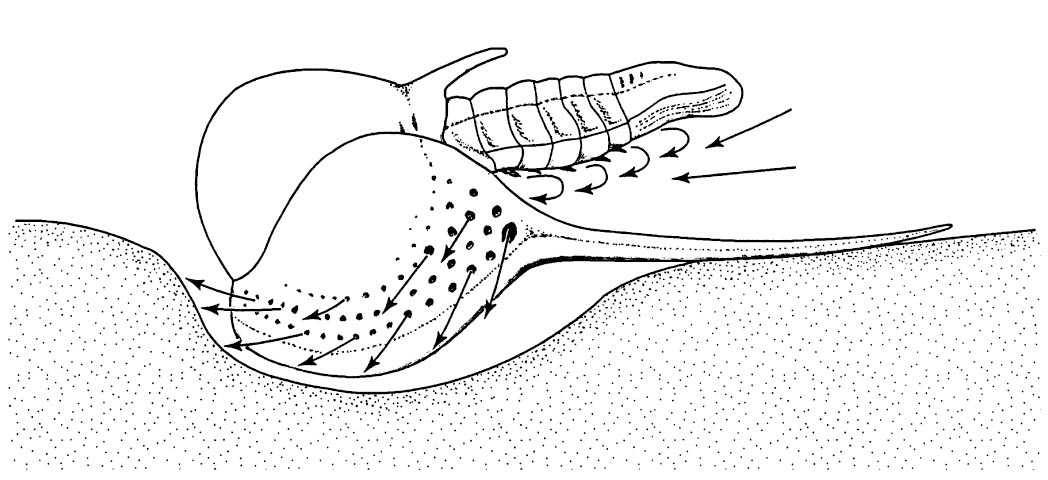
Diagram outlining the trinucleid ‘chamber filter-feeding’ process, as first hypothesised by Fortey and Owens 1999.

This digging of sediment was suggested to be a possible cause of Rusophycus; a common trace fossil, long assigned to trilobites as a resting trace. Copyright free image.

Whereas Rusophycus is formed by a stationary trilobite, Cruziana is thought to represent a burrowing/furrowing trace, as the trilobite hunted/scavenged through the sediment. Cruziana will often start with a Rusophycus, demonstrating the two can be produced by the same animal exhibiting different behaviours. Copyright free image.
Unless otherwise stated, all images courtesy and copyright of Peter Cameron. All rights reserved.
Related Posts

Episode 174: A History of Dinosaurs in 50 Fossils →

Episode 102: Small Shelly Fossils →

Episode 94: Joggins Fossil Cliffs →











