
Evidence of Intense Predation Pressures on Ancient Megafauna
Biology is full of exciting avenues, and some of the finest, in my opinion, are the morphological and behavioural adaptations that define the split seconds whether an animal lives or dies, eats or starves. Predator-prey interactions are extremely exciting- not only visually (I strongly recommend watching the BBC’s The Hunt)- but they also play an important role in the dynamics of biological systems.
Some of you may remember the announcement of an exquisitely preserved nodosauid that made the news a few months back, including a spread in the June edition of National Geographic. Nodosaurids are a clade within the ankylosauria, famed for their armour, although they differ from their ankylosaurid cousins most notably via the lack of an obvious tail club, preferring a more flexible caudal series. Amazingly, the Suncor nodosaur, as it is more colloquially know, is so well preserved that not only are the overlying osteoderms still in life position, but soft tissue was fossilized as well. And importantly, yesterday marked the online release of its official name, as well as exciting information regarding its ecology and soft tissue preservation.

Fig. 1. Borealopelta markmitchelli, the new nodosaurid. Note the lightly coloured parascapular spines and the light underbelly, compared to the reddish brown dorsal integument. Credit: Julius Csotonyi/Courtesy of the Royal Tyrrell Museum of Palaeontology, Drumheller, Canada.
Northern Shield
Caleb Brown and colleagues present Borealopelta markmitchelli (Fig. 1), a rather typical nodosaurids, nestled in the middle of the phylogeny, weighing in at 1300kg and measuring approximately 5.5.m in length (unfortunately the tail is missing, preventing a more accurate size estimation). What is atypical about the specimen is where the carcass was deposited: at the bottom of the sea. The rock formation that housed Borealopelta for 110 million years is more famous for its marine reptiles that its dinosaurs, the nodosaur being the first of its kind. Unsurprisingly, the carcass drifted to the bottom of the sea on its back, most likely weighed down by all that dermal armour.
Two aspects of this fossil make it stand out from the ankylosaur crowd: the presence of melanin in the large parascapular spine, integumentary scales and the epicuticle between scales, preserved as an organic, keratin-derived film. Melanin is one of the more common pigments in vertebrates and comes in three basic types: eumelanin (responsible for black and brown colours), pheomelanin (which produces red), and neuromelanin, although the latter is exclusive to the brain.
Sociosexual Spines
The parascapular spine, located unsurprisingly in the shoulder region, was shown to possess a keratin sheath that was lighter in colour, compared to the remaining sheathes, in visible light, and slightly fluorescent under UV light. This is indicative of a reduced melanin concentration in this structure, and thus would have been lighter in colour in life. These standout spines, not only in colour but also size, may have acted as a display structure, promoting fitness or age to their conspecifics. Importantly, the authors note, this evidence bolsters the idea that dermal armour in ankylosaurs was not only an anti-predator adaptation, but acted as a plastic, multifunctional tool for several behaviours. The use of parascapular spines as a sociosexual display unit could possibly be extended to some of the stegosaurs, the structure being common in the basal forms like Gigantospinosaurus, whilst also cropping up in the more derived Kentrosaurus. It would be interesting to see if a certain sex possessed a different coloured parascapular spine to the other, but that is wishful thinking at best.
The Influence of Megapredators on Herbivore Colouration
Areas of preserved skin, when subjected to pyrolysis-gas chromatography-mass spectroscopy (py-GC-MS for short…), noted the presence of benzothiazole, diagnostic of pheomelanin, responsible for reddish hues. The extent of this pigmentation can be seen in a cross section of the sacrum, with a darker dorsal surface transitioning to a less pigmented ventral surface. This sort of pigment distribution is akin to countershading in extant vertebrates, and has important implications for an animal’s ecology.
Countershading, also known as Thyer’s law, is a form of crypsis, and is denoted by a variation in the dorsal and ventral colouration of an animal’s body, with darker colours on top and vice versa on the belly. The physics behind this colouration, and one of the several theories for its function, pertains to how light falls on a three dimensional object. Imagine an evenly shaded animal: light from the sun falls over it’s back, making it lighter, whilst the shade generated by the animals bulk makes the ventral aspect darker. As a result, the animal looks solid, giving its presence away. The ability to incorporate pigmentation where it matters reduces this effect, helping the animal remain unnoticed. This is not unique to herbivores, predators need to conceal themselves too if they are to effectively stalk their prey.
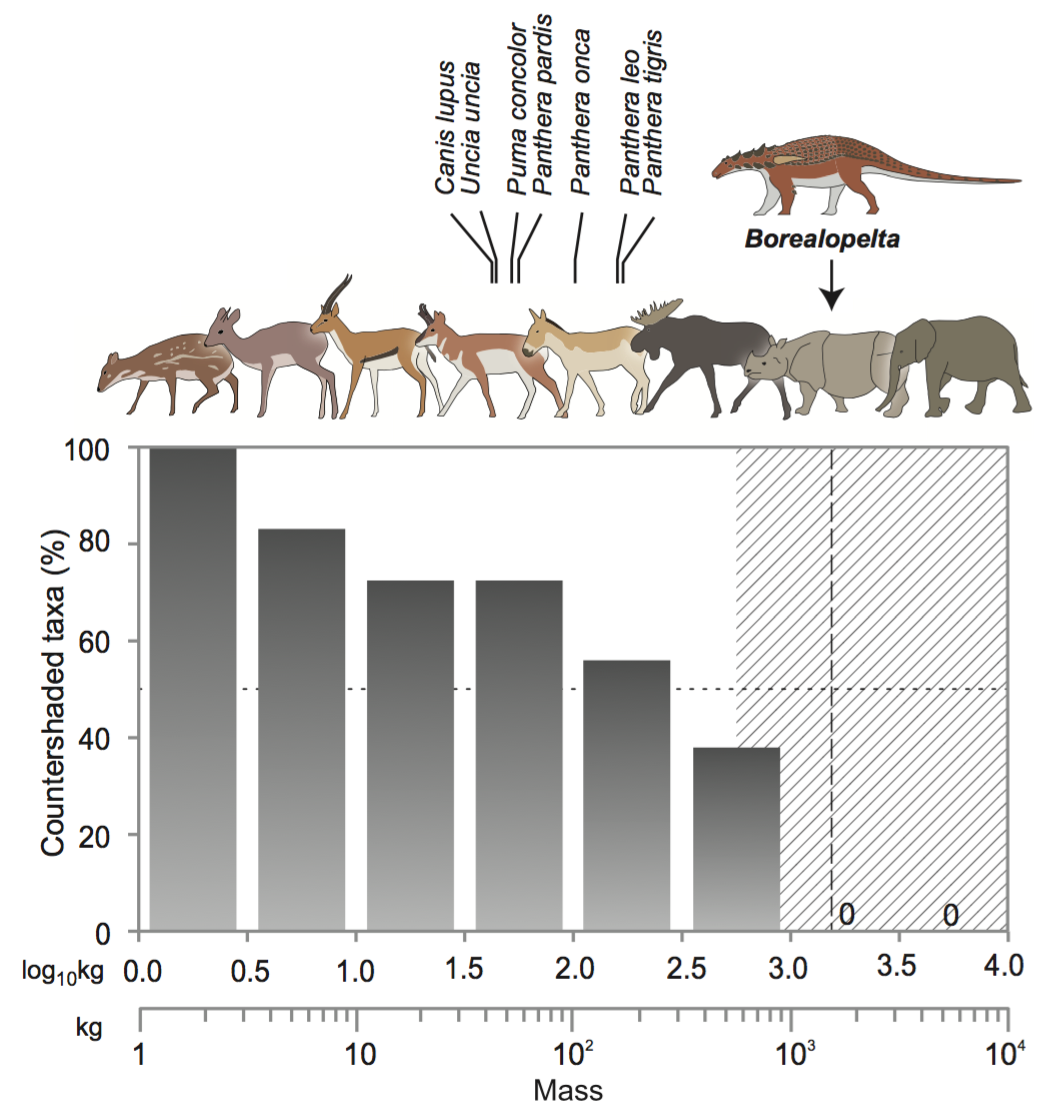
Fig. 2. Graph showing the loss of countershading in larger animals, with largest predators labelled in italics. Credit: Brown et al., 2017.
What is of particular interest here is the fact that we’re seeing countershading in an extremely large terrestrial herbivore. If we look at Brown et al.’s (2017) analysis of modern mammalian assemblages, we can see a cut off point whereby animals get so big they no longer need any form of crypsis; their size is enough to deter predation attempts (Fig. 2).
This isn’t the first time countershading has been noted in dinosaurs, following the extraordinary Psittacosaurus specimen showing preserved pigmentation (Fig. 3). In the little ceratopsian, the countershading was positioned low on the body, indicative of a “closed” light environment, for instance a forest. Animals in more direct sunlight, in open environments like a plain, tend to possess a gradient higher up on the body. The very ventral position of the countershading gradient in Borealopelta could hint at a similar “closed” environment.
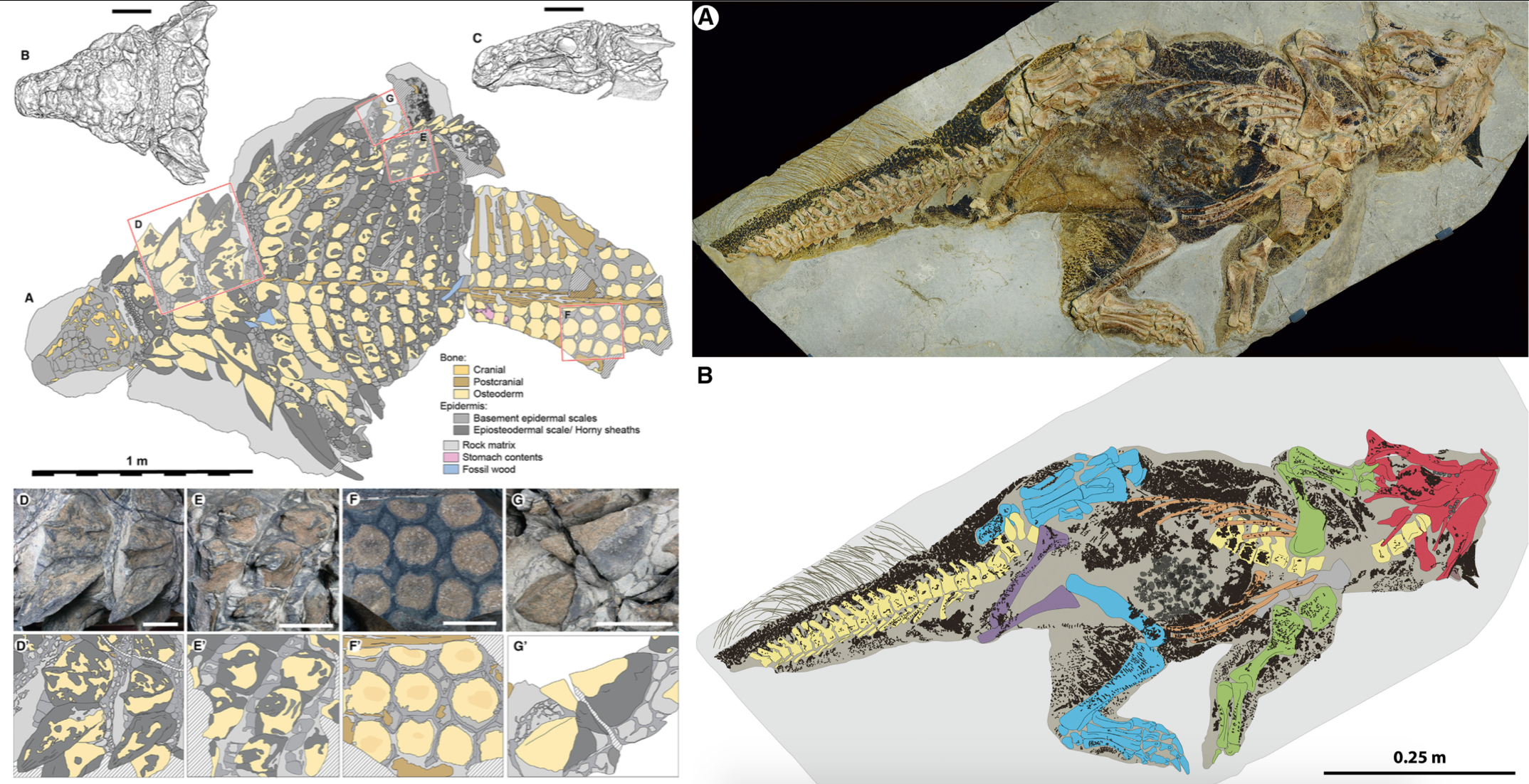
Fig. 3. Distribution of soft tissue in Borealopelta (left) and Psittacosaurus (right). Credit: Brown et al., 2017; Vinther et al., 2016.
Unlike the aforementioned mammalian ecosystems whereby other mammals prey on smaller individuals, promoting the evolution of countershading in theses species, Mesozoic ecosystems were devoid of anything remotely like the large extant carnivorans at the apex of food chains; that honour went to the large theropods. Unlike the dichromatic mammals (possessing two colour receptors-cone cells- in the retina), theropods would have been at least trichromatic, if not tetrachromatic, like modern day birds. This, combined with the excellent vision seen in crocodilians, the other half the dinosaur extant phylogenetic bracket, has the authors feeling that the need to hide from large, visually acute predators would have been of the utmost importance, even in particularly large, well armoured thyreophorans like Borealopelta. This, of course, is unlike anything in modern ecosystems today. As we have already mentioned, Borealopelta was discovered in marine sediments, so there is no accompanying evidence of large theropods associated with the animal. However if we look at members of geological formations of similar age and locality, we will notice the presence of huge carcharodontosaurid: Acrocanthosaurus.
Mistaken Identity?
Interestingly, writing in the aforementioned Psittacosaurus article, Jakob Vinther and his team speculatively suggested that predators of the little ceratopsian may have used shape-from-shading cues to identify prey (hence the need to evolve countershading). However, I wonder whether predation pressure from Acrocanthosaurus, or similar sized theropods, was the cause of countershading in the herbivore. Acrocanthosaurus was a tall predator, and as such would have been looking at prey from less acute angles compared to smaller predators, which would have looked at Borealopelta effectively in lateral view (Fig. 4), rather than variations in dorsal view. As far as I can tell, countershading in terrestrial vertebrate herbivores is due to predation pressures from animals that stalk their prey low to the ground, i.e. canids and felids. In these animals, the head is rarely raised during the stalk, and as a result view their prey side on. This provides the best image of a self-shading object, causing the necessary need to co-evolve countershading measures.
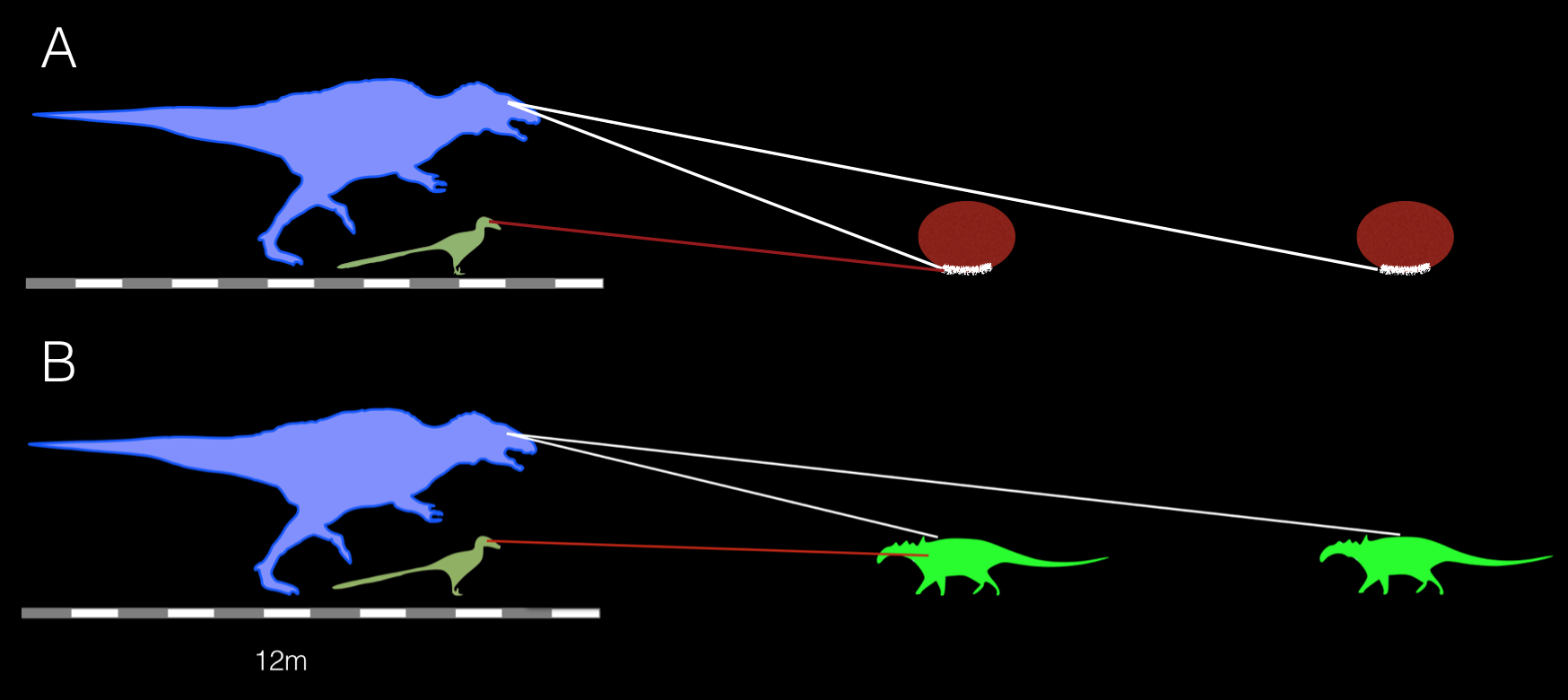
Fig. 4. A very unrigorous schematic representing the lines of sight of Acrocanthosaurus and Deinonychus, compared to a Borealopelta at varying distances, viewing the herbivore in lateral (A) and frontal (B) view. The larger predator has a predominately dorsal view of the nodosaurid, and would not have seen much of the countershading (white line under the red circles, based on the extent of melanin in the cross section of the sacral region(Brown et al., 2017- Supplementary materials), whereas the smaller maniraptoran is able to see the animal in a more lateral view. This, coupled with the poor carcharodontosaurian binocular vision at low elevations, means I feel it is unlikely Acrocanthosaurus would have placed selection pressures for countershading. Credit: debivort, Dinoguy2/wikicommons; Chris Barker.
I’m arguing that it is not an Acrocanthosaurus-type predator that is placing predation pressures on Borealopelta to develop countershading, but a smaller theropod. One of the other sparse theropods from the region and time was the maniraptoran Deinonychus, roughly half the height of an adult human. Not only are they found in a clade known to possess enhanced optic lobes, they also have greater binocular fields compared to carcharodontosaurids like Acrocanthosaurus, providing better depth perception. The tall rostrum of of Acrocanthosaurus would have limited it’s binocular field of view, especially at low elevations, most likely faring similarly to its cousin Carcharodontosaurus in that respect, perhaps employing panoramic surveillance instead. Ultimately, as Vinther and colleagues (2016) write, “it is not well known how widely shape-from-shading is utilized in the animal kingdom outside of humans“, and as such we can only speculate on this image perception in predatory theropod dinosaurs.
Furthermore, are these predators really big game specialists? Derived tyrannosaurs are the pinnacle of big game hunting, using extreme osteophagy to get away with attacking large prey. But the same couldn’t be argued for the two theropods mentioned above, even despite the size of Acrocanthosaurus, and I’m not aware of any evidence of predation on ankylosaurs. In addition, the presence of unarmored iguanodontians in rocks of the same age and locality must have surely served as better prey items. This is of course speculation on my part, but I would be interested to find out more from someone with better knowledge of vision in predatory animals, and the influence of shape-from-shading cues in prey detection. Also, a more accurate and scientifically backed diagram denoting the effect of height, binocular field of view and countershading wouldn’t go amiss either…
The presence of those lightly coloured parascapular spines is also interesting, as one could argue they stand out on the animal, perhaps defeating the point of countershading. Maybe they were meant to conspicuous, to deter predation attempts via a visual signal of fitness, or perhaps they were employed as a form of disruptive colouration, helping bolster the countershading. Such patterning further breaks up an animal’s three-dimensional shape, reducing the chance of detection due to the contrasting colours, which are often matched by the surrounding environment. This is unlikely though: in lateral view, the spines would present themselves as a small, shaded, roughly ovoid shape, surely not enough to disrupt the overall image of the animal. Furthermore, the effect of osteoderm-generated shade has yet to be seen, and would be interesting to analyse its effect on countershading.
The predation pressure hypothesis for countershading in large herbivorous dinosaurs is an exciting one, and one that deserves further study to elucidate whether this was actually the case. It could be the colours we are seeing are vestiges of juvenile colouration, where countershading would have been more important due to their small size and lack of cursoriality, or perhaps even an attractor for the opposite sex. Nevertheless, this fine fossil has delivered an excellent publication, and one that I hope spurs further work into the subject area.












