Life, molecules and the geological record
To understand the evolution of life, palaeontologists can employ a variety of techniques. This typically involves the visual identification of fossils, like bones or teeth, within the sedimentary record, either by eye or using a microscope. However, life also leaves molecular fossils within the geological record.
Molecular fossils (or “biomarkers”) are organic molecules which can be tied to a specific biological source and include nucleic acids (e.g. DNA and RNA), proteins, carbohydrates and lipids. Of these, lipids are the most highly resistant to degradation and can be preserved over millions of years. As such, they can be used to investigate the evolution of life and past ecosystem dynamics.
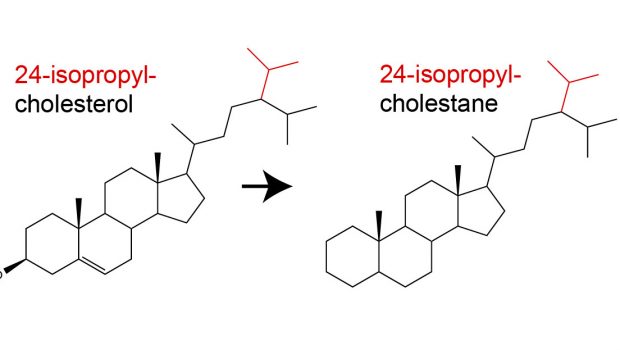
One class of lipid molecules which are particularly informative are sterols. These lipids are almost exclusively made by eukaryotes and are involved in a range of cellular functions (e.g. maintaining membrane fluidity and stability). Within the geological record, there are a diverse range of sterol structures, one of which is known as 24-isopropyl-cholestane (24-ipc). This curious compound is a diagenetic product of 24-isopropyl-cholesterol (Fig. 1a), a sterol found within certain genera of demosponges (Figure 1b). As such, it has proposed as a marker for fossil sponges within the geological record.
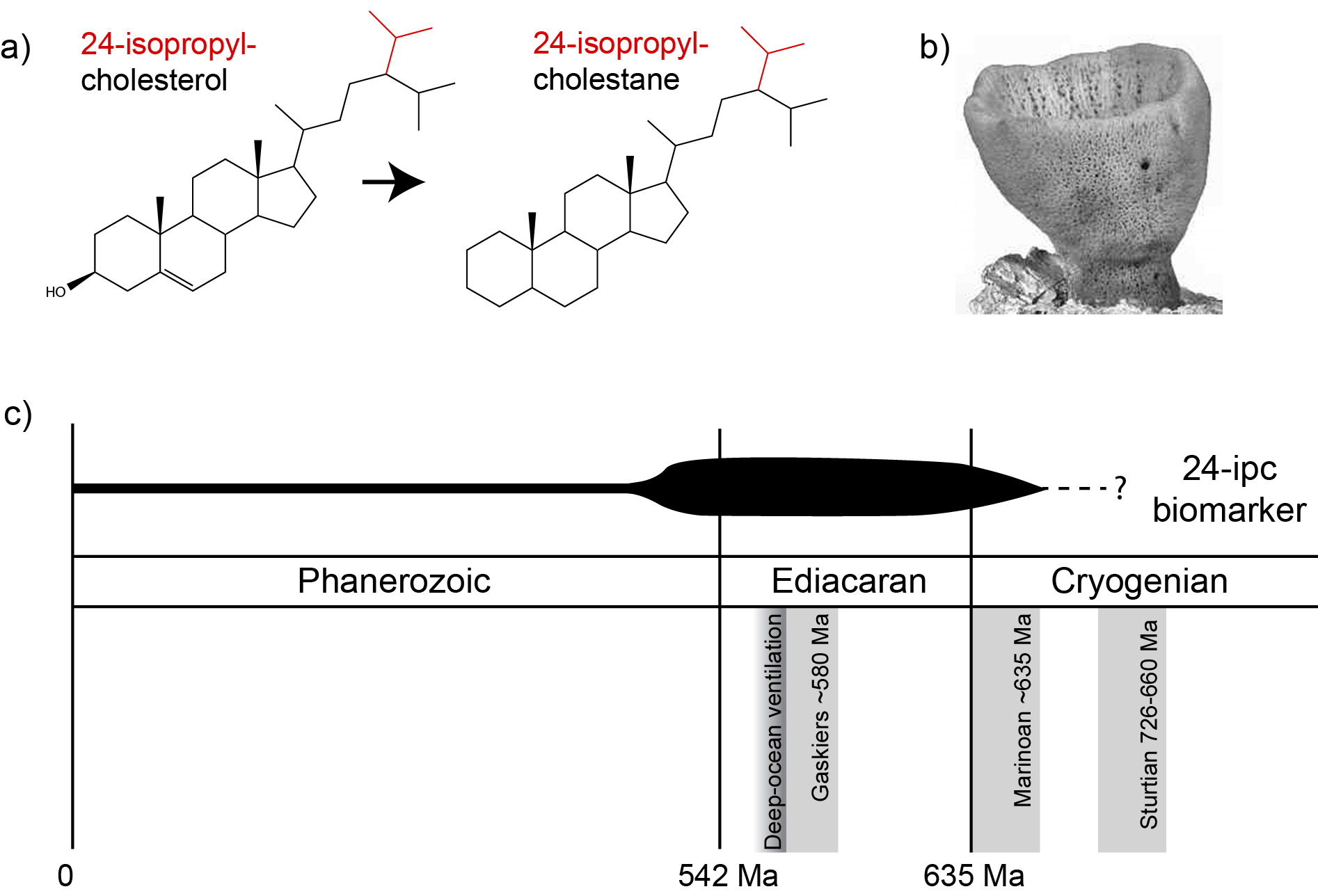
In 2009, researchers at the University of California Riverside and MIT observed the elevated presence of 24-ipc within ~640 million year old sediments (Fig. 1c; Love et al., 2009). This finding was remarkable and provided evidence for complex animal life up to 100 million years prior to the Cambrian explosion. This was also much earlier than the existing fossil evidence for demosponges (sponge spicules; ca. 550 Myr ago). Despite these findings, some researchers have questioned a sponge origin for this biomarker (Antcliffe, 2013; see reply by Love et al., 2013). For example, it is known that other organisms (namely, pelagophyte algae) can also produce 24-ipc. Despite this, recent genetic studies argue that Demospongiae acquired the ability to synthesise 24-ipc up to 100 Ma before this trait was acquired by pelagophyte algae (Gold et al., 2016). As such, this compound likely remains the earliest evidence for complex animal life in the geological record.
Another important class of lipids are highly branched isoprenoids (Fig. 2a). These lipids are made by diatoms, unicellular algae which are important primary producers in today’s ocean (Fig. 2b). HBIs are notable for the distinctive “T-branch” in their carbon skeleton (Fig 2a, in red) and for having up to six double bonds. Highly branched isoprenoids are also relatively resistant to degradation and have been used to explore the rise of diatoms during the Phanerozoic (Sinninghe Damsté et al., 2004) Analysis of marine sediments and petroleum extracts revealed that C25 HBIs first appeared in the fossil record during the upper Turonian (91.5 ±1.5 Myr ago; Fig. 2c). This predates the first physical evidence for diatom fossils by ~20 Ma and has been attributed to relatively poor preservation of physical diatom fossils, which are often prone to dissolution.
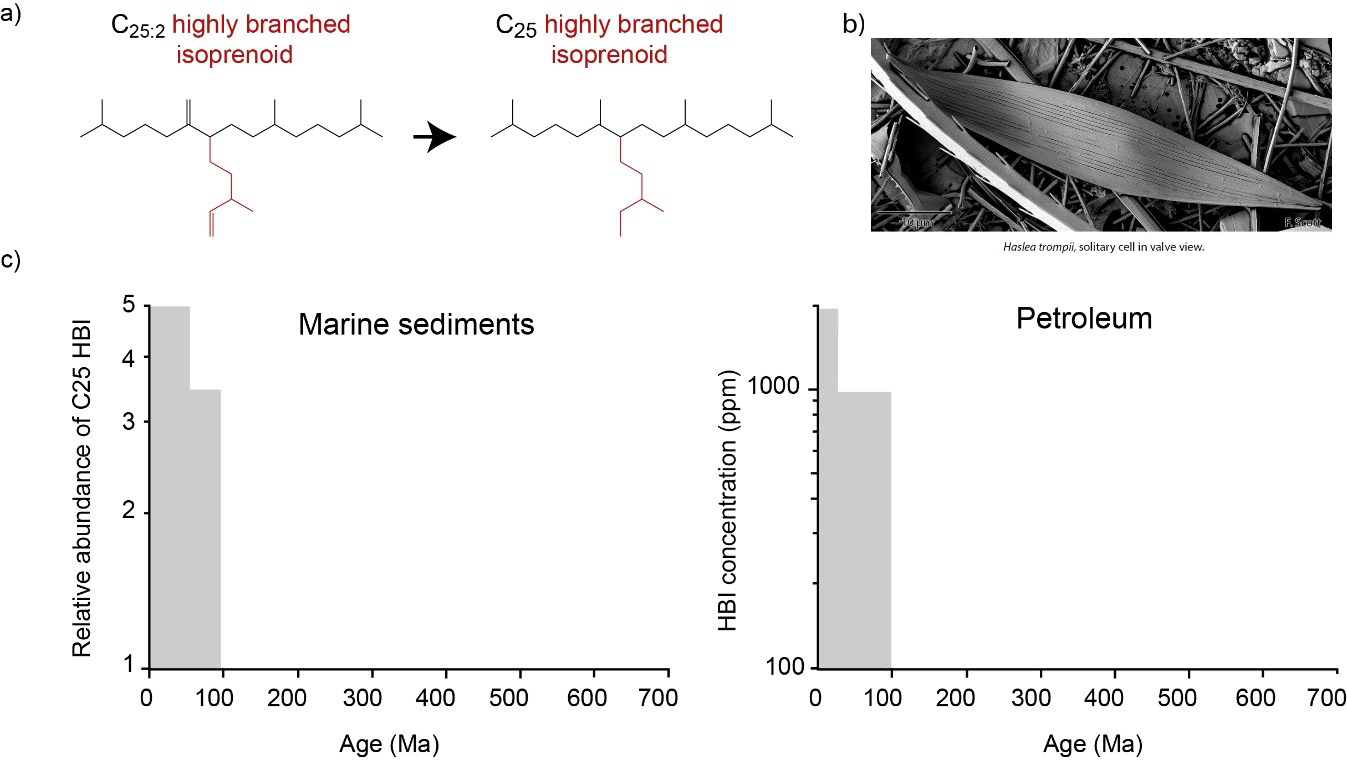
Figure 2: Highly branched isoprenoids in the geological record: a) degradation of C25:2 HBI to C25 HBI, b) the modern diatom (Haslea trompii), c) relative abundance of C25 HBIs within sediments (left) and petroleum (right) over time (redrawn from Sinninghe-Damsté 2004)
Biomarkers can also be used to study the evolution of life on land. One of the earliest fossil molecules studied was also one of the simplest; the n-alkanes (Eglinton and Hamilton, 1967). These are straight-chained hydrocarbons (Fig. 3a) which form part of the waxy, protective outer coating of plant leaves. Plants have two main ways to capture solar energy; C3 and C4 photosynthesis. The former is present in most grasses, trees and shrubs (i.e. the majority of plants). The latter is less common, but is used by several economically key plants, including maize, sugar cane and millet. Crucially, the differences in these two pathways (C3 vs C4) results in distinct stable carbon isotopic signatures (Fig. 3b; Cerling et al., 1997). The advent of the C4 photosynthetic pathway is a relatively recent geological innovation; however, due to the poor preservation potential of grasses, it remains unclear when this originated.
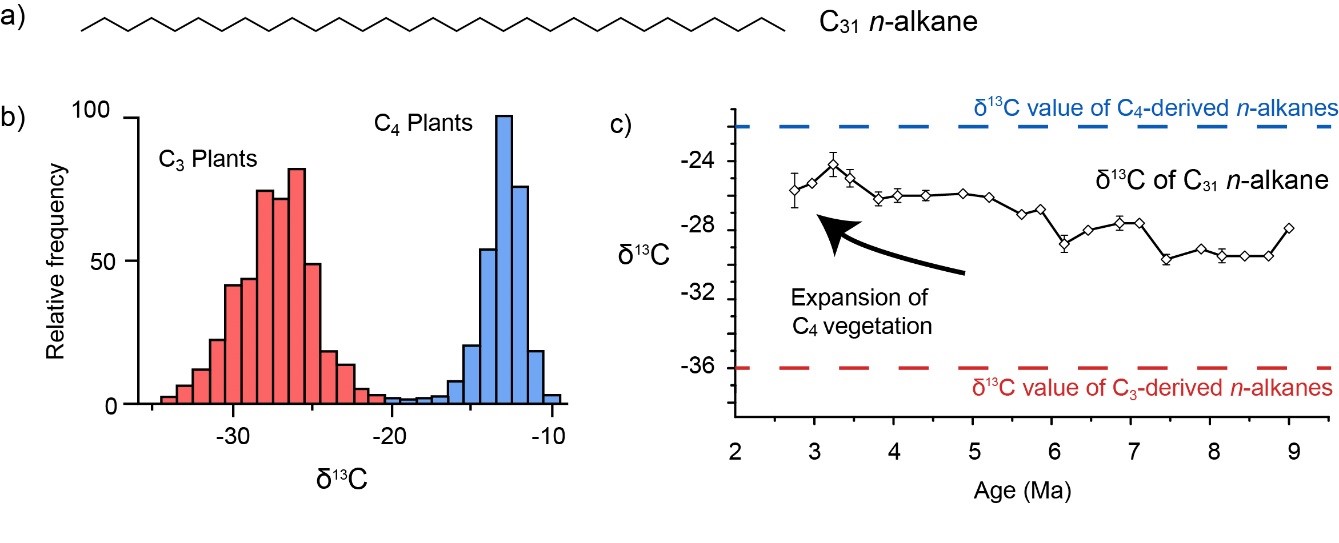
Figure 3: Tracing the onset of C4 photosynthesis. a) C31 n-alkane in both its biological and geological form, b) the carbon isotopic composition of C3 and C4 plants (redrawn from Cerling et al., 1997), c) input of C4 vegetation during the late Miocene and early Pliocene (redrawn from Hoetzel et al,. 2013)
One way to explore this is to analyse the carbon isotopic composition of n-alkanes. Generally, the δ13C value of long-chain n-alkanes in C3 savannah vegetation averages −36‰ (Vogts et al., 2009). In contrast, those for C4 grasses average around −22‰ (Vogts et al., 2009). This approach was successfully applied to sediments from offshore Namibia, where the C31 n-alkane indicates a long-term positive carbon isotopic excursion during the late Miocene and early Pliocene (Fig. 3c). This suggests greater contribution from C4 vegetation and is consistent with other approaches (e.g. tooth enamel). Expansion of C4 grasslands during the late Miocene coincides with declining CO2 concentrations, increasing aridity and decreasing temperatures (see Herbert et al., 2016) and suggests that C4 photosynthesis was developed in order to cope with certain environmental pressures.
Collectively, these studies showcase the utility of molecular fossils. Due to their ubiquity and preservation potential, they can be used to reconstruct the evolution life over different geological timescales and have become an essential component of the modern palaeontologist’s toolkit.
Glossary:
Alkane: synonym for saturated hydrocarbon (i.e. a chemical compound that contains only carbon and hydrogen, and has no double bonds or aromatic units).
Biomarker: an organic compound that can be unambiguously linked to specific precursor molecules made by living organism
C3 & C4 photosynthesis: two different metabolic pathways for carbon fixation during photosynthesis. The two pathways produce different amounts of carbon isotope fractionation, such that the biomarkers can be distinguished by their δ13C values.
δ13C: A measure of the relative amount of the two stable carbon isotopes in a substance.
Isopropyl: an alkyl group wherein the three carbon atoms are arranged in a Y shape
References:
Antcliffe, 2013. Questioning the evidence of organic compounds called sponge biomarkers, 56, 917-925
Brocks and Butterfield, 2009. Biogeochemistry: Early animals out in the cold. Nature, 457, 672-673
Cerling et al., 1993. Expansion of C4 ecosystems as an indicator of global ecological change in the late Miocene, Nature, 361, 344-345
Cerling et al., 1997. Global vegetation change through the Miocene/Pliocene boundary. Nature. 389. 153-158
Eglinton and Hamilton, 1967. Leaf epicuticular waxes, Science, 156, 1322-1335
Gold et al., 2016. Sterol and genomic analyses validate the sponge biomarker hypothesis. PNAS. 113, 2684-2689
Herbert et al., 2016. Late Miocene global cooling and the rise of modern ecosystems. Nature Geoscience. 9, 843-847
Love et al., 2009; Fossil steroids record the appearance of Demospongiae during the Cryogenian period, Nature, 457, 718-721
Love et al., 2013. The molecular record of Cryogenian sponges –a response to Antcliffe (2013), Palaeontology, 58, 1131-1136
Sinninghe Damsté et al., 2004. The rise of the Rhizosolenoid Diatoms, Science, 304, 584-587
Vogts, et al., 2009. Distribution patterns and stable carbon isotopic composition of alkanes and alkan-1-ols from plant waxes of African rain forest and savanna C3 species. Organic Geochemistry, 40, 1037–1054
 Description of author:
Description of author:
Gordon Inglis is a Postdoctoral Research Associate working within the Organic Geochemistry Unit at the University of Bristol. He is an organic geochemist and palaeoclimatologist who uses biomarkers to reconstruct temperature, hydrology and biogeochemistry during the early Cenozoic.
@climategordon http://gordoninglis.strikingly.com/
Related Posts

Episode 171: Freshwater Mosasaurs →

Episode 153: Castle Bank →

Episode 123: Soil →











