Oxygen Isotopes and Oviraptorosaurs
Implications for Dinosaur Nesting Behaviour and Thermophysiology
Oviraptorosaurs had received a pretty bad reputation when they first popped onto the scene. Initially, palaeontologists branded these creatures as “egg plunderers”, due to the discovery of an individual, Oviraptor, atop of a pile of eggs that were thought to belong to Protoceratops, a sheep-sized, desert-dwelling ceratopsian. Henry Osborn, describing the animal in 1924, gave the supposed egg-thieving theropod its ignominious name, although he did note that despite appearing to be caught red handed, it’s name “may entirely mislead us as to its feeding habits and belie its character”.
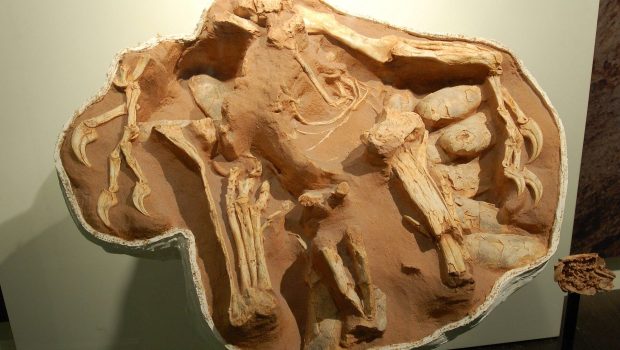
And right he was. More recent (and fantastic) discoveries have shown these dinosaurs possessing brooding habits similar to that of birds. Oviraptor wasn’t trying to steal the eggs; it was caring for them. The wonderful fossil of Citipati, a cousin of the accused Oviraptor, shows the animal sitting in the middle of it’s nest, its arms draped over the double layer of eggs that form the inner circumference of the mound (Fig. 1). Amazingly, embryonic individuals are known to science, still curled up in their eggs, undisturbed for over 80 million years.

Fig. 1. Citipati osmolskae, brooding its clutch. Credit: Dinoguy2/Wikimedia.
The fact that these animals sit on their nests implies that they must be doing so for temperature related reasons, for instance to provide warmth or shade for their developing young. A recent paper by a French-Chinese research team sought to find out what temperature these eggs were brooded at, using oxygen isotopes derived from the calcium carbonates of eggshells and phosphates from embryo bone.
Bear with me as we go through the importance of oxygen isotopes…
The utility of oxygen isotopes is well documented in palaeontology. Spinosaurs had their ratios measured from the enamel of their teeth to reveal a semiaquatic ecology, revealing evidence of niche portioning. Media darling Tyrannosaurus and its equally large cousin Giganotosaurus had their oxygen isotope ratios measured from their dorsal vertebrae, ribs and legs, with results suggesting a constant core temperature regardless of seasonal air temperature variations. Importantly, conclusions from isotopic oxygen ratios suggest these animals were endothermic and homeothermic, capable of generating and maintaining elevated body temperatures within a known mammalian/avian range.
Naturally occurring oxygen comes in three stable forms: 16O, 17O and 18O, the former being the most abundant (by about 99.8%), and also the heaviest. Due to different weights, these isotopes can be fractionated, producing a ratio, and importantly, the degree of fractionation is dependent on temperature: warmer temperatures result in more 16O.
Thus, as a vertebrate embryo develops and its bones mineralise, it incorporates these isotopes by stripping the eggshell of its calcite (calcium carbonate) content and depositing them as a phosphate. As a result, this incorporation process would thus reflect the egg temperature and the oxygen isotope composition of the water surrounding the embryo, denoted as d18Oew (originally derived from their calcitic shell, d18Oc).
However, several factors affect the oxygen isotope composition of bone phosphate (d18Op), including water vapour loss via the eggshell pores to embryo metabolism, which prompted the researchers to produce and propose a physiological model in order to predict the changes in embryo bone d18Op values versus various incubation temperatures and at changing d18Oew., based on known fractionation factors and physiological traits of extant archosaurs.
Odd Oviraptorosaurs
If you thought last weeks post on abelisaurs mentioned bizarre anatomy, oviraptorosaurs are possibly even weirder. Their snouts are reduced to a large, beak-like jaw, which is usually toothless in the more derived forms (although some primitive individuals of the group have goofy, prominent buckteeth…). Some had crests adorning their skulls, and they had begun to reduce the number of caudal vertebrae in their tails. Feathers have been found on several species, with a fan of them possibly adorning the reduced tails of some, and anchor points on the ulna of others suggest feathers were attached to the arms. The latter is significant, as these feathered forelimbs would perfectly envelop a brood of eggs, as suggested by the position of the arms in the fossilized remains of doting parents (see Fig. 1).
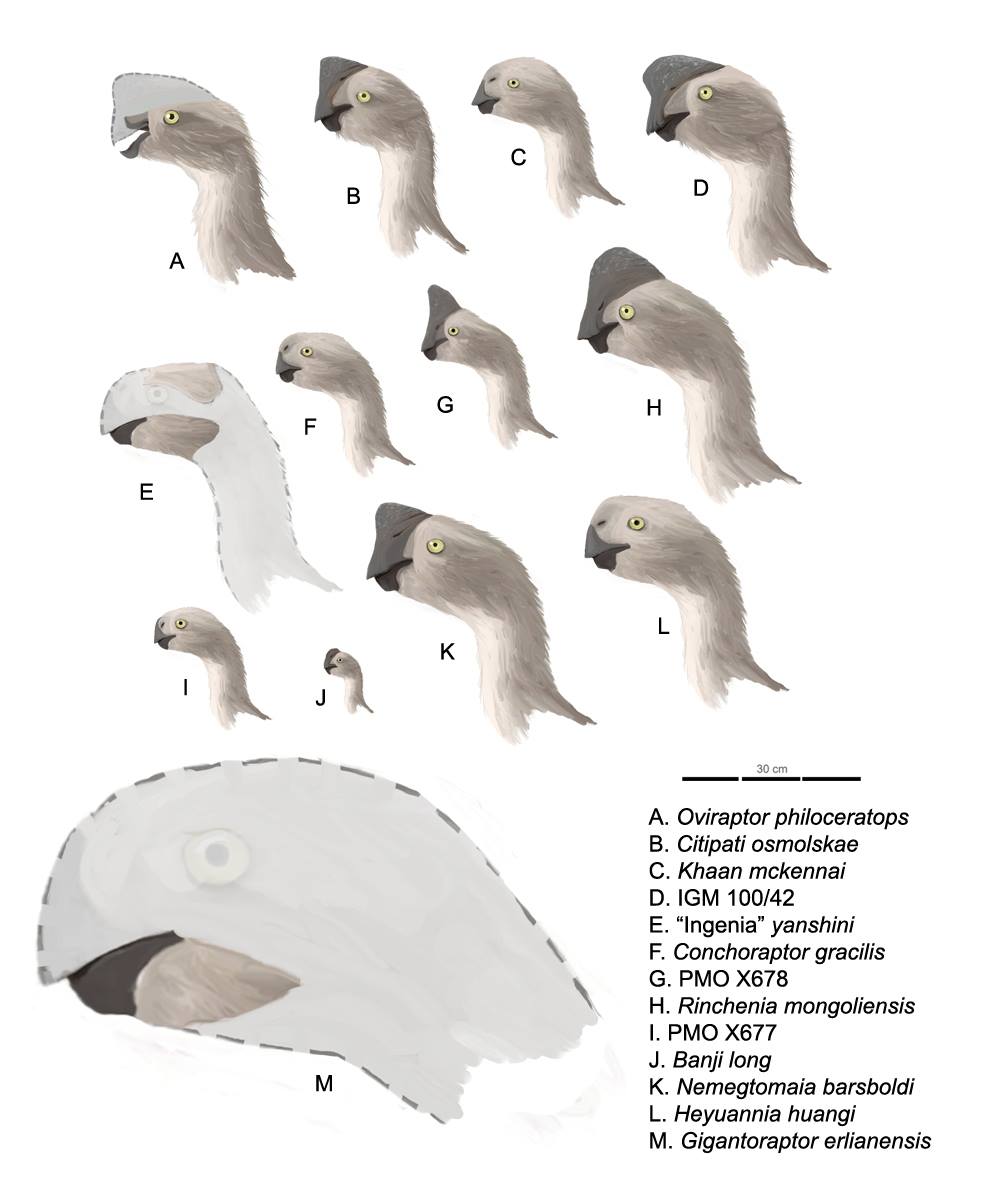
Figure. 2. Oviraptorosaur skull profiles. Note the enormous size of Gigantoraptor and crested heads of some its cousins. Credit: Dinoguy2/Wikimedia.
Eggshells and Endothermy
Eggshell calcite and bone phosphate were measured from seven elongatoolithid eggs (i.e. a family of eggs assigned to oviraptorosaurs) from the Upper Cretaceous of China, all of which contained embryonic skeletons, as were two Nile crocodile eggs (in order to complement the known data for oxygen isotope ratios of eggshell and egg water in modern archosaurs).
Assuming constant incubation temperature, one would expect the bone phosphate d18Op value of the embryo to also reflect the oxygen isotope composition of the mother’s body fluids, and as a result a linear relationship should form between d18Op and d18Oc. However, science isn’t perfect and likes to ruin your dataset just for laughs, with five of the eggs clustering at weirdly high d18Oc values. The modeled incubation temperature that would fit both d18Oc and d18Op values indicated these individuals would have had to have been incubated at temperatures over 45 degrees, which is not only beyond any known modern species, but is also a lethal temperature for many eggs, especially over a sustained period. The possible culprit, according to the authors, is additional carbonate from an external source, most likely during the fossilisation period (although, to add to the anomaly, microscopy of those specimens revealed little recrystalisation…).
The isotopic oxygen analyses revealed the rest of the eggs were incubated within 35-40 degrees centigrade, with incubation estimated at 44-56 days, similar to results suggested by Lee (2016) for the aforementioned Oviraptor and Citipati. Interestingly, these results are far shorter than those predicted by a recent study by Erickson and colleagues (2017) on ornithischian taxa, which are estimated to require three to six months. The authors suggest two possible explanations: the parameters used in Lee’s 2016 data are derived from extant birds, which may have shortened the incubation time due to their derived physiology, or that the ornithischians in Erickson et al.’s paper retained the ancestral reptile state, whilst the theropods begun adopting the rapidity seen in their modern day bird descendants. Whether the latter explanation holds up against sauropod data, or even the proposed shake up of the dinosaur family tree, remains to be seen.
Interestingly, the estimated bone formation temperature is higher than those estimated via clumped isotope analysis for oviraptorosaur eggs from Mongolia. Eagles and colleagues presented results in 2015 showing formation temperatures of around 31.9 degrees centigrade. The authors note this is peculiar, especially considering the fact that this clade is close to the avian branch of the family tree (with some even suggesting they are primitive birds). As always, more research is needed to figure out the cause of this discrepancy, although the authors note that their model could be applied to other egg-laying amniotes, extinct or extant, that “could provide new insight into the environmental constraints that influenced the geographical distribution of biodiversity in terms of reproductive strategies and capabilities”.
 Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Related Posts

Why did Sauropods grow so big? →

200 Years of Dinosaurs →

Episode 158: Ceoptera evansae →













