
Neovenator Neurovasculature and the Purpose of Sensitive Snouts
Articles on dinosaur neurovasculature are like buses: you wait for most of the Phanerozoic for one to arrive and then two appear at once. Indeed, 2017 saw the publication of two articles relating to the purpose of cephalic neurovasculature in theropod dinosaurs, the subject being only briefly highlighted in a select few cases from disparate Mesozoic reptiles. Thanks to micro-focus computed tomography (μCT), our discovery of complex internal canals within the rostrum (snout) of Neovenator salerii, a midsize carnivorous theropod from the Isle of Wight, is the most complete to date, and broke the trend whereby such canals were thought to facilitate aquatic foraging in fossil reptiles.
Prior to our results, CT imagery brought out internal canals in the premaxilla of the giant marine reptile Pliosaurus, as well as in rostrum of the large African-and seemingly semiaquatic-theropod Spinosaurus. Both of these creatures were compared to extant crocodilians, who share similar neurovasculature complexes and surface foramina on the bones of the snout. We wanted to investigate what was behind the foramina on the bones of Neovenator, which had a plethora of them across their lateral surface.
In modern archosaurs, foramina interact with the surrounding skin to form integumentary sensory organs (ISOs). One use of this organ in crocodilians is to improve aquatic foraging, capable of detecting prey generated ripples in the water. As such, it was thought that both Pliosaurus and Spinosaurus employed a similar mechanism to detect prey in an aquatic/semiaquatic environment. Baryonyx, also a spinosaurid, is said to possess a similar complex. We wanted to see whether this would be the case for Neovenator too.
In addition, along with our Neovenator article, a second terrestrial theropod was thought to possess a sensitive snout following bone surface analysis of the newly named Daspletosaurus horneri. I will comment further on the this individual a bit later on, however the implications of these two papers suggests that the phylogenetic bracket for complex rostral neurovasculature has expanded to no longer solely include spinosaurids, but may include tetanuran theropods as a whole.
What is a Neovenator?
Neovenator salerii is one of the largest carnivorous dinosaurs discovered in the UK, and yet it remains rather poorly known. This theropod dinosaur discovered on the Isle of Wight in the 1970s, from a band of rock known collectively as the Wealden Group, dates back to the Barremian age of the Lower Cretaceous, approximately 125 million years ago. When it was described Neovenator was thought to be the oldest European allosaurid, although this has since been refuted with the discovery of older allosauroids including Torvosaurus and Allosaurus. The placement of Neovenator within allosauroids has been debated, however more recent phylogenetic analysis firmly places N. salerii as the basal-most individual within the Carcharodontosauria, forming its own clade along with several other “problematic” theropods: the Neovenatoridae. Our individual, the holotype, is a subadult, and appears to share the typical hallmarks of an allosauroid, occupying the niche as a terrestrial predator. It is 70% complete, suggesting a svelte, relatively nimble animal, most likely preying on hypsilophodontids and iguanodontids that roamed the Early Cretaceous.
A Brief Overview of Rostral Neurovasculature Structure
The cephalic neurovasculature in Neovenator is beautifully preserved, becoming increasingly branched as the complex moves anteriorly through the maxilla and into the premaxilla, terminating on the surface of the bone via small neurovascular foramina. Like the Spinosaurus scan results, it would appear that there is no interaction with any of the sinuses in the skull. Interestingly, the complex is laterally situated in comparison to the tooth row, leading us to believe that it had little influence on the teeth itself (one could expect that any dental use for such a structure would surround the teeth, rather than being placed on the labial side of the alveoli). Furthermore, a thermoregulatory function, as seen in modern crocodilians and some birds (REFS) via a palatal plexus, appears unlikely as there was no indication the complex innervated medially, nor is there a large palate to support such a structure. In addition, the narrow skull of Neovenator is taller that it is wide, reducing the area for palatal heat exchange, and the anterior position of the majority of the neurovasculature branches further reduces the area for heat exchange to occur. Thus, a sensory function seem the most plausible use for this structure.
We identified these canals as containing branches of the trigeminal nerve, the fifth cranial nerve (CN V), which splits into three sections: the ophthalmic branch (V1), the maxillary branch (V2) and the mandibular branch (V3). The largest of the cranial nerves, it is responsible for motor and sensory information being passed between the jaws and brain. In Neovenator, as in many vertebrates, the ophthalmic branch relayed sensory information from the premaxilla, whilst the maxillary branch did the same in the maxilla. The dentary was not available to us, however the neurovascular foramina that dot its surface, particularly anteriorly, leads me to believe that a similar complex is mirrored in this lower jaw bone (see fig. 2 in Brusatte et al, 2012).
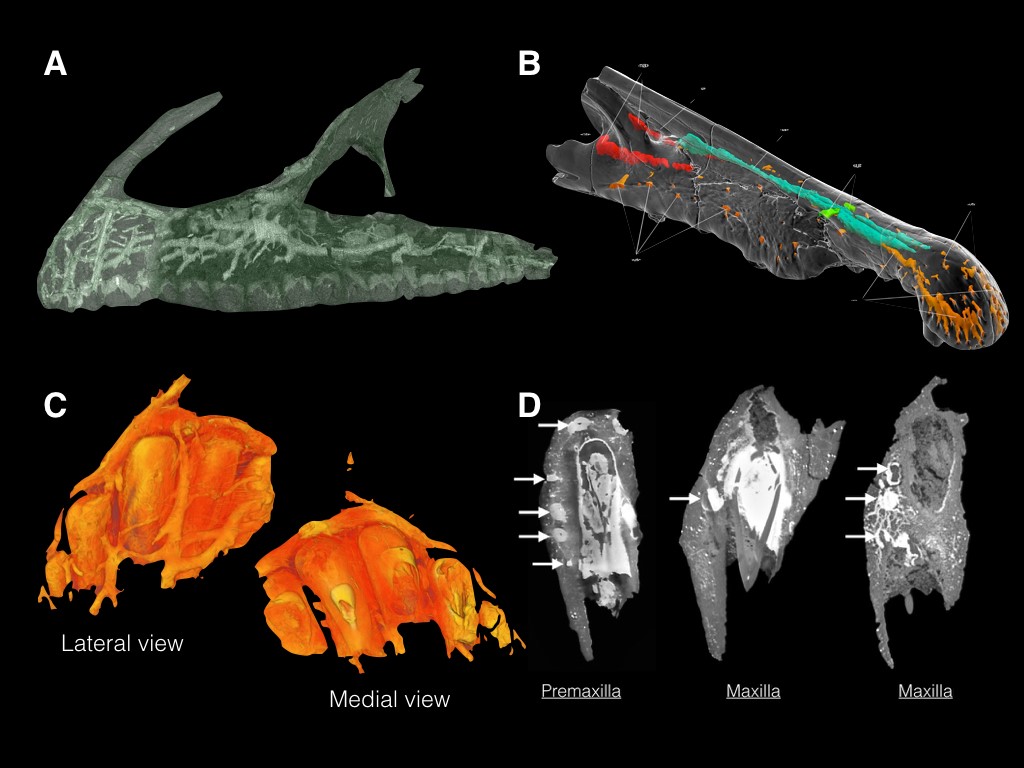
Figure. 1. A) Complex neurovasculature in the snout of Neovenator; showing almost complete preservation; B) CT rendering showing the partial preservation of rostral neurovasculature in Spinosaurus; C) CT rendering of the Neovenator left premaxilla, the bone at the tip of the snout, showing the branched pattern over the lateral side of the bone, and it’s absence on the medial side; D) Transverse slices of the premaxilla and maxilla of Neovenator, further illustrating the lateral placement of the neurovascular complex. Images not to scale. (Image credits: Barker et al., 2017/Chris Barker, Ibrahim et al., 2014, Fig. S6)
What’s the Use of Facial Sensitivity in a Mindless Killing Machine?
As previously discussed, enhanced facial sensitivity is seen in crocodilians, as well as several clades of bird, including ostriches, corvids, parrots, several shorebirds, and even the humble kiwi. But what use is it in a 7 metre long theropod predator? Surely it doesn’t care what it’s doing with its snout? Our first port of call came about following scanning electron micrographs of its teeth generated as part of my Masters thesis. In hindsight, 3D textural analysis, the superior cousin of 2D SEM imagery, may have provided more quantitative results, nevertheless the images produced from several of its teeth seemed to suggest that Neovenator did not like bone, in fact, it seemed to be actively avoiding it. Comparisons with SEM studies on modern day mammalian carnivores- (admittedly not the best clade to contrast with) showed a scratch-dominated enamel surface texture, with very minimal pitting. This is akin to cheetahs, that actively avoid bone, preferring to feed on the flesh of its prey. We proposed that Neovenator was doing something similar: deliberately avoiding tooth-bone contact thanks to the sensitivity of its snout. The ability to differentiate between bone and meat may have proven useful for a creature that did not utilise bone matter in the same way tyrannosaurids do.
However ISOs are capable of more than just pressure reception. Chemical cues and thermoreception have been noted in animals with these sensory organs. Whilst detection of chemicals in the air is mainly done in the nose (and in some organisms the specialised vomeronasal organ), thermorecepetion may have been more pertinent to theropods like Neovenator. It would appear that the more basal theropod dinosaurs created nest mounds out of vegetation, much like todays crocodilians and some megapode birds (also known as incubator birds). In both of these clades, eggs can be deposited in a large mound of vegetation, which as it rots and ferments, produces heat in which to incubate the eggs. Both the crocodilians and megapodes, which provide a degree of parental care, are capable of adjusting the temperature of the nest using their rostra. It is perhaps not unfeasible to suggest that theropod dinosaurs followed similar behaviour, with Carr et al. (2017) even suggesting they used their sensitive snouts to delicately move offspring in the way mother crocodiles do.
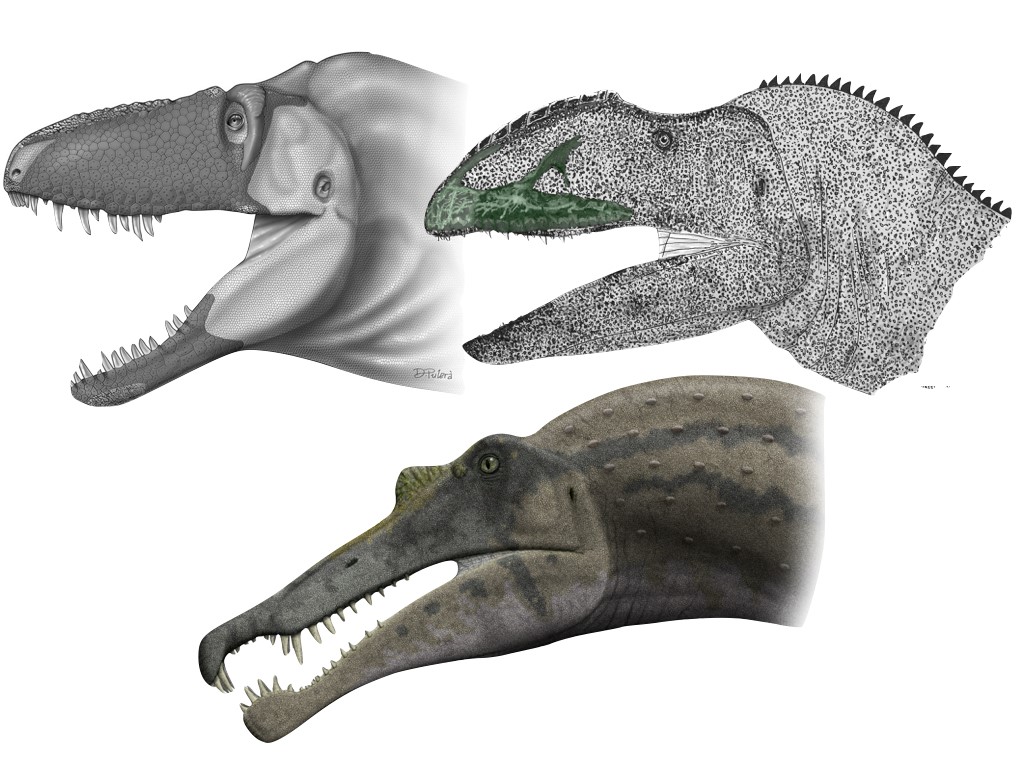
Figure. 2. Some theropods with inferred snout sensitivity… From left to right: Daspletosaurus horneri (credit: Carr et al. 2017/Dino Pulerà), Neovenator salerii (credit: Barker et al., 2017/Darren Naish), and Spinosaurus aeygptiacus (credit: wikicommons/Steveoc 86)
The Tyrannosaur Conundrum
As mentioned above, earlier this year Carr et al. (2017) published an article on the aforementioned newly identified Daspeltosaurus horneri, a large tyrannosaur from Montana, United States. They came to similar conclusions regarding theropod facial sensitivity, however lacked the CT data seen in our Neovenator specimen. Interestingly, they suggested that the facial skin was covered in large flat scales (the terminology used is incorrect, crocodiles don’t have facial scales, but rather a heavily cracked integument), where the neurovascular foramen are densest, based on comparisons with modern alligators and several bird species. Myself and some of my coauthors were intrigued by this conclusion, due to two notable aspects of tyrannosaur biology. Firstly, the method of prey dispatch and feeding is unique in derived tyrannosaurs whereby the skulls are designed to withstand extremely large forces and deliver bone crushing bites (the most recent models indicate a 4 tonne bite force in T. rex coupled with over 400, 000 psi at the tooth tip!), and seem to have produced more bite marks than other theropods. Furthermore, facial wounds are common in tyrannosaurs, indicative of intraspecies aggression, whilst only occurring in select cases amongst other theropods.
It would be of interest, therefore, to explore:
- The prevalence of facial wounds in crocodilians, in order to assess whether the prevalence of facial injuries and enhanced sensitivity can be linked (i.e. do crocodilians avoid getting head wounds to protect their ISOs?);
- The size of the internal neurovasculature in tyrannosaurs compared to animals like Neovenator and Spinosaurus, as well as extant archosaurs with tactile snouts, taking into account the effects of allometry and ontogeny, as well assess whether canal volume can infer environmental/behavioural cues in extinct and extant organisms;
- And to assess whether differences exist in the volume of the trigeminal fossa, the area of the brain that produces the trigeminal nerve branch, in animals with enhanced rostral sensitivity compared to those without.
There is much more work to be done in this field, but unfortunately for myself and my colleagues, we lack the resources to explore this field of science further. Nevertheless, we have plans in place to delve into dinosaur (and even archosaur) facial neurovasculature in the future, with the hope to unravel what could be a very important aspect of dinosaur physiology and behaviour.
 Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Blog post by Chris Barker. Chris studied Vertebrate Palaeontology at the University of Southampton and has a fondness for all things archosaur.
Related Posts

Episode 174: A History of Dinosaurs in 50 Fossils →

Episode 171: Freshwater Mosasaurs →

Society of Vertebrate Paleontology Annual Meeting 2025 →











