
New Oviraptorid Shows Cassowary Convergence
The Late Cretaceous rocks of Ganzhou, China, are rife with oviraptorids. We have seen these strange theropods before here at Palaeocast, when we looked at the very high temperatures at which they incubated their eggs. The Ganzhou rocks, which range from Campanian to Maastrichtian in age, have six oviraptorid species, and their ranks have been bolstered by a new addition: Corythoraptor jacobsi.

Fig. 1. Artist representation of Corythoraptor jacobsi, with a clear nod to modern cassowaries not only in the crest but also the colouration of the facial integument. Credit: Zhao Chuang/Lü et al. 2017.
Junchang Lü and colleagues noted several diagnostic characters that separate it from the other Ganzhou oviraptorids, and were particularly impressed by the cranial ornamentation in this species, which greatly resembled a cassowary’s crest. Bone histology of the rib, right fibula and left radius were analysed in an attempt to estimate age, with the fibular growth marks suggesting the animal was at least eight years old. It appears that it was yet to reach full body size, and died as a sub adult at the beginning of a new season, as seen by the deposition of new bone close to the periosteal surface (an outer bone surface).
Cassowary Conundrum
Cassowaries are one of my favourite birds, mainly due to their dinosaurian appearance, cool head and neck colouration, and the fact they can beat most people up. These birds, native to New Guinea, some neighbouring islands, and parts of Australia, possess a distinctive, tall crest on their heads, found in both sexes. The utility and function has remained a mystery, however my friend and colleague Darren Naish published an interesting paper on this peculiar bit of anatomy (as well as on his blog, Tetrapod Zoology, which is always worth a read). Inside the leathery, keratin sheath is an assortment of densely packed trabeculae (thin bony struts) and air spaces, similar to what was found in the Corythoraptor holotype specimen. (Fig. 2.)
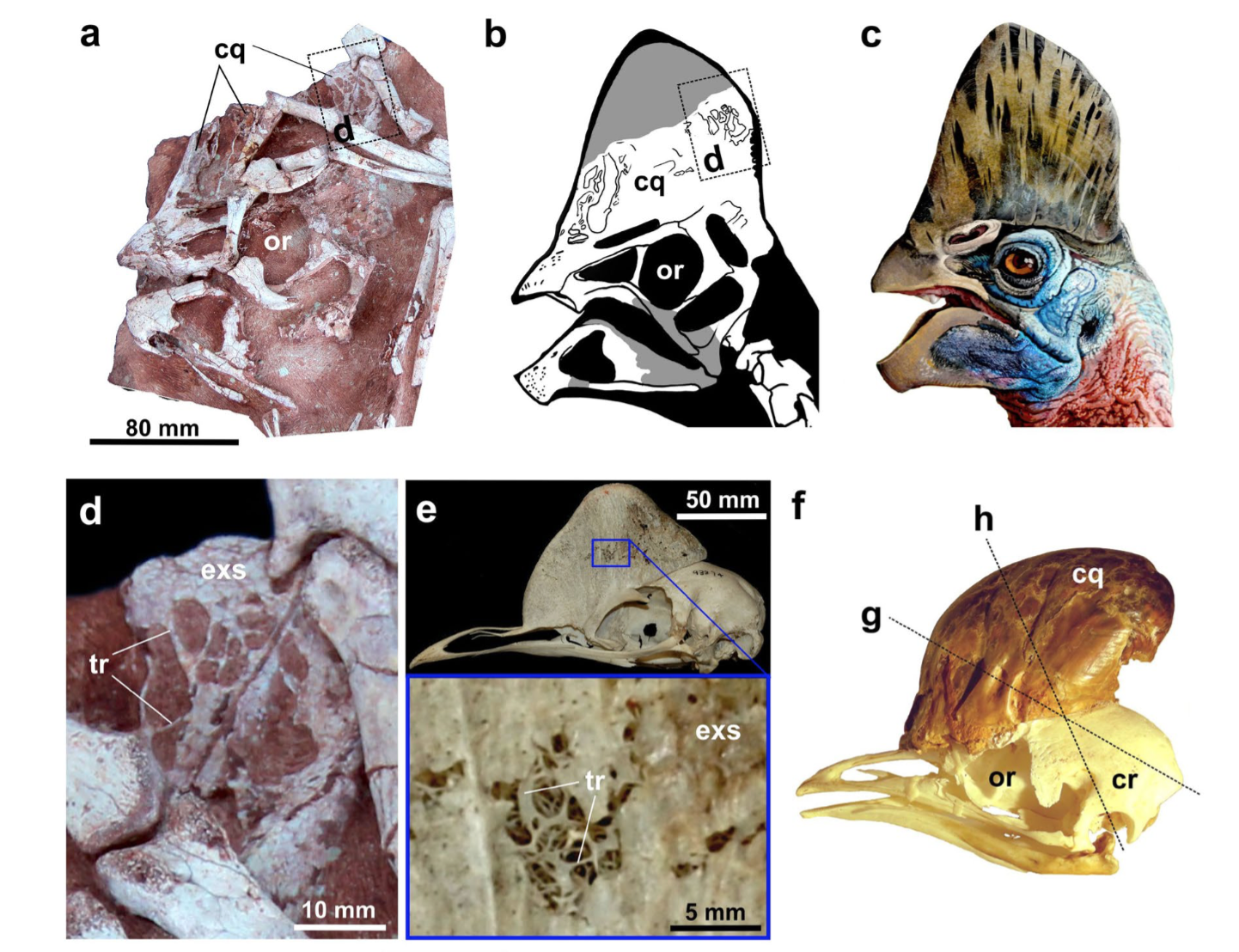
Fig. 2. (a-c) Skull of Corythoraptor, showing the tall casque and internal trabeculae,(d), not unlike that of the modern cassowary (e-f). Credit: Lü et al., 2017.
As Darren notes, most hypotheses relating to function are anecdotal, and some are rather bizarre, but the most likely use related to visual sexual display, which coevolved as a resonating chamber. Cassowaries make the deepest call in the avian kingdom, and the crest is angled as such to direct these vocalisations towards a prospective partner. Interestingly hearing capabilities are known for some crested oviraptorids, such as Conchoraptor, which are tailored to the low frequency range (thanks to pneumatic pathways in the skull).
However, cassowary calls are generated in the throat, and Corythoraptor appears to lack the morphology to be capable of this, and as such it is improbable the casque was used solely in an acoustic fashion. Intraspecific visual signals of fitness and hierarchy are more probable, and presence of this crest in a sub adult implies that Corythoraptor was sexually active prior to reaching adult size. Prey capture and predator avoidance could also play a role, particularly if the crest acted as a resonator, especially with large, weird-looking tyrannosaurids patrolling the area.
The presence of this crest in both cassowary sexes suggests mutual sexual selection, and whether this would be applicable to Corythoraptor is of interest, but unfortunately sexing extinct reptiles is particularly hard, and analysis of medullary bone, present in ovulating females, is unreliable. A final note relating to function is the use of the crest in cassowary thermoregulation. As Lü notes, there is evidence that the crest is used to help shed heat in the extant bird, which could have evolved a similar function in this new oviraptorid. It is an old paper, and I would like to see an updated CT-based project to explore this function in greater depth (I’m looking at you, WitmerLab!).
Why there are so many oviraptorids in Ganzhou is unclear, but Lü, writing in 2016 on Corythoraptor’s cousin Tongtianglong, offers two explanations. The first, known as taxonomic over-inflation, could be down to the fact that there has been a glut of recent discoveries in the area, with some of these newly described individuals perhaps representing previously described species. Differences in sex (sexual dimorphism) or development (ontogeny), or even random variation could result in seemingly differing morphologies in animals of the same species (e.g. the difference in tadpoles and adult frogs, for instance, or the sexual display of the male peacocks versus their drab female counterparts). And whilst this is unlikely, with all seven oviraptorid species possessing species-specific traits (autapomorphies), the large difference in size between some of the species, as well as the potential for developmental changes occurring in the cranial ornaments (as seen in the horned ceratopsids and crested hadrosaurs), could point to an ontogenetic bias.
The more likely scenario suggests there was a genuine diversification event of oviraptorids in the end Cretaceous rocks of the Nanxiong Formation of Ganzhou. And whilst ontogeny most likely influenced crest shape and sixe in oviraptorids, very few diagnostic characters are actually based on this part of the skull. The huge variation in other cranial elements, such as the bones of the jaw and associated musculature, suggests variability in diet, which usually remains conserved throughout life. This is best explained by species separation, perhaps via food-related niche partioning.
Adding to the mix is the fact the Ganzhou rocks are poorly dated. It may be that these oviraptorids actually lived over a large time span and were not sharing a similar locality, and were instead separated by thousands or millions of years, which is being obscured by our current lack of knowledge of the geology of the area.
Related Posts

Episode 174: A History of Dinosaurs in 50 Fossils →

Episode 171: Freshwater Mosasaurs →

Society of Vertebrate Paleontology Annual Meeting 2025 →











